© Е.Г. Зеленова, М.И. Заславская, Т.В.Махрова, 2002 г.
УДК 616.992
Поступила 9.04.2001 г.
Е.Г. Зеленова, М.И. Заславская, Т.В.Махрова
Государственная медицинская академия, Нижний Новгород
Кандиды: экология, морфофункциональные особенности и факторы патогенности
Впервые упоминание о кандидозных поражениях (инфекциях) ротовой полости было обнаружено у Гиппократа в IV в. до н.э. (377 г. до н.э.). Местные описания кандидозных поражений сделаны в 1771 и 1784 гг. Сandida аlbicans впервые была выделена от больного туберкулезом в 1844 г., в 1849 г. дрожжеподобные грибы обнаружены при вагинальном кандидозе, а в 1853 г. – при системной инфекции.
В XX в. серьезное исследование микроскопических грибов рода Саndida было начато в Австралии. В 1939 г. на 3-м Международном конгрессе микробиологов австралийский исследователь доктор Вергхоут предложил принять термин Сandida для возбудителей микозов. Официальное имя рода Саndida было утверждено на 8-м ботаническом конгрессе в Париже в 1954 г. 1.
Этиология
Термин «молочница» (monilia) впервые был использован ботаниками при описании поражения овощей в 1751 г. 2. Как заболевание человека молочница названа Гиппократом stomata aphtoidea, а Галленом – aphta alba infantu. Перевод на русский язык греческого слова кандида означает «кандидат в белой мантии (одежде)» (в древней Греции сенаторы одевались в белые одежды, что указывало на чистоту и непорочность). Саndida albicans – дважды повторенный эпитет «белый» в видовом названии основного представителя кандид (лат. albicans – белый). По-видимому, исследователи, предложившие это название, хотели акцентировать внимание на специфических местных проявлениях, которые возникают при кандидозах.
Классификация кандид
Род Сandida включает около 150 видов грибов, которые относят к дейтеромицетам в связи с полным отсутствием половой стадии развития. Некоторые (семь) из этих видов: С.albicans, C.tropicalis, C.krusei, C.kefyr, C.(Torulopsis) globrata, C.guilliermondii, C.parapsilosis – признаны с медицинской точки зрения наиболее важными болезнетворными микроорганизмами 3, 4. C.albicans является наиболее распространенным представителем рода, изолированным от человека, и известен как комменсал и как условно-патогенный микроорганизм 1, 5
Экология кандид
Различные виды Сandida обитают в разнообразных экологических нишах. Все эти дрожжеподобные грибы широко распространены в природе. Термин «дрожжи», применительно к таксономии грибов, не является официальным. Его чаще всего применяют к грибам семейства Saccharomycetaceae (класс Endomycetes, порядок Ascomycota), а под «дрожжеподобными» организмами понимают формы, размножающиеся преимущественно почкованием клеток, не подразумевая при этом какого-либо таксономического родства между ними 6. Так, из более чем 2000 генов, идентифицированных у С.albicans, только 100 (5%) имеют молекулярную гомологию с Saccharomyces 7.
Дрожжи отличаются от плесневых грибов (плесеней) по двум признакам: во-первых, колонии дрожжей на питательных средах похожи на бактериальные – гладкие, пастообразные, вырастают через 48-72 ч; во-вторых – при микроскопировании дрожжей выявляются клетки овальной или круглой формы, разной величины (процесс почкования); размеры клеток – до 10 мкм. Такие же морфологические и культуральные признаки имеются у кандид.
Кандиды могут находиться на объектах внешней среды, на поверхности медицинского инструментария, на предметах домашнего обихода, на игрушках, посуде, в питательном креме для рук, косметике. Они лучше выживают на влажных поверхностях, чем на сухих неживых объектах, но при достаточной степени загрязнения могут сохраняться на последних до 24 ч 8. С.albicans изолируется с зубных щеток большинства носителей кандид во рту, c зубных протезов, катетеров. В больших количествах эти микроскопические грибы обнаруживаются в молочных продуктах: в творожных сырках (78,6%), твороге (66,6%), сметане (75%). Кандиды способны сапрофитировать в окружающей среде на субстратах живой и неживой природы (чаще на богатых сахарами фруктах и овощах, особенно несвежих) 2, 9.
С. albicans выделяется от приматов, домашних и других млекопитающих, сумчатых, птиц 5. У людей С.albicans предпочтительно колонизирует поверхность слизистых (ротоглотка, влагалище). Желудочно-кишечный тракт является главным резервуаром инфекции, при этом С.albicans способна колонизировать практически любую часть желудочно-кишечного тракта, от ротовой полости до перианальных тканей 10, 11. Вульвовагинальная область колонизирована видами кандид у 40% здоровых женщин 5. С.albicans также часто колонизирует человеческий эпидермис, особенно влажную кожу между пальцами рук или ног. Частота выделения кандид от людей зависит от возраста и диеты: ротовая полость – 1,9-41%, желудочно-кишечный тракт – 0-55%, влагалище – 2,5-68% 4.
При носительстве в ротовой полости первое место занимает С.аlbicans – 47-75%, второе – С.tropicalis и С.globrata – 7% 3. К факторам, которые увеличивают процент носительства кандид, относят: снижение процесса слюноотделения, низкую рН слюны, увеличение концентрации глюкозы в слюне, курение. Отмечено также, что носительство С.аlbicans у пациентов в стационаре значительно выше, чем у здоровых людей. Так, частота орального носительства у здоровых людей – 17%, у больных в стационарах (без клинических проявлений кандидоза) – 40-69%. До сих пор окончательно не доказано: постоянные или разнообразные штаммы могут присутствовать у людей при непрерывном и длительном носительстве. Заметим, что постоянное выделение С.аlbicans из полости рта еще не является доказательством кандидозной инфекции (см. ниже).
Возбудителем кандидоза чаще всего является С.аlbicans (до 62% случаев) 4. У иммунокомпрометированных хозяев кандидозные инфекции поддерживаются резидентными штаммами 12, 13. Эпидемиологический анализ с использованием отпечатков ДНК биотипов показал, что большая часть штаммов, ответственных за инфицирование, генетически сходны 14, 15. Из внешней среды С.аlbicans выделяется реже, чем другие виды кандид, и это может быть следствием ее большей патогенности (адаптация к паразитизму) для человека 1.
Состояние здоровья каждого индивидуума является предрасполагающим фактором для колонизации С.аlbicans. Проявление заболевания может зависеть от колонизируемой ткани, факторов вирулентности клеток кандид и ответной реакции хозяина. В свою очередь колонизация зависит от нескольких факторов:
приобретения или попадания кандид в полость рта или другие экологические ниши;
адгезии и собственно колонизации;
проникновения в ткани;
удаления кандид с поверхности слизистых (механизмы клиренса) 5.
Предрасполагающими факторами для возникновения орального кандидоза и патологии, ассоциированной с ним, являются 3:
возраст (неонатальный и пожилой);
пищевой дефицит;
антибиотики;
опухоли;
ВИЧ-инфекция;
химиотерапия;
кортикостероиды (гормонотерапия);
носительство зубных протезов.
Передача кандид в клинических стационарах (нозокомиальный кандидоз) возможна при контакте с носителями, через руки персонала, путем контаминации при поцелуях (в 2 мл слюны может быть от 300 до 500 клеток кандид) 16. Так как кандиды обнаруживаются в обработанных и сырых продуктах, есть возможность для занесения их в ротовую полость руками, через слюну, контаминированную пищу или питье 17.
Попадание клеток кандид на слизистые оболочки не является достаточным для колонизации и инфекции 3, 5. Оральная полость является экологической нишей с постоянным перемещением ротовой жидкости, поэтому дрожжевые клетки, если они не адгезированы и не размножаются, могут быть вымыты слюной или проглочены. Удаление кандид со слизистых может обеспечиваться и иными механизмами: перистальтические сокращения ускоряют прохождение микроорганизмов через желудочно-кишечный тракт, а поток мочи поддерживает стерильность мочевого пузыря 18.
Морфология кандид
Существуют различные морфологические варианты кандид: от дрожжей до гифальных форм. Гифальные элементы считаются более агрессивными и патогенными, в то время как дрожжеподобная форма является комменсальной и менее патогенной 10. Хотя кандиды часто называют диморфными, фактически они являются полиморфными, так как могут принимать вид дрожжей, гиф (несептированный мицелий) и псевдогиф (псевдомицелий) – тонких удлиненных клеток, располагающихся друг за другом в виде нитей и не имеющих общей оболочки 1, 5, 6, 9.
Для кандид характерно только неполовое размножение, при этом формирующиеся в процессе жизнедеятельности неполовые споры принято называть «конидии» (греч. konis – пыль). С. аlbicans могут образовывать гроздеподобные скопления бластоконидий (молодые клетки гриба диаметром 2-5 мкм). Бластические конидии кандид отпочковываются от материнской клетки и располагаются на перетяжках псевдомицелия.
Для С.аlbicans характерно формирование ростовых трубок, предшествующее возникновению новых гиф и дрожжевых форм 1, 2, 6. Так, если С. аlbicans инкубировать 2 ч при температуре 37оС в 10% сыворотке, то из бластоконидий формируются типичные удлиненные клетки – ростовые трубки. Они отличаются от других подобных выростов отсутствием перетяжек на основной трубке (гифе). Одним из факторов, стимулирующих образование ростовых трубок (и далее гиф) служит повышенное содержание углекислого газа (с такими условиями микроскопические грибы сталкиваются в тканях).
Отличительной особенностью С. аlbicans является также образование хламидоконидий (спор с двойной плотной оболочкой) на концах или коротких боковых отростках гиф. Хламидоспоры служат для переживания неблагоприятных условий 1, 2, 6, 9, 19.
Отмечено, что при поверхностном кандидозе кожи и слизистых в очагах поражения преобладают бластоспоры, а при глубоких микозах обнаруживаются в основном вегетирующие клетки 19.
Температура оказывает важное влияние на морфогенез кандид. Так, температура около 25оС способствует формированию хламидоконидий. Более высокая – от 37 до 40оС (подобный диапазон температур может быть в организме у потенциальных больных) и реже до 43оС – содействует образованию гиф и псевдогиф 20. Однако не все виды кандид способны расти при 37оС и выше, поэтому данное свойство является важной патогенетической характеристикой и отделяет потенциально патогенные штаммы от сапрофитов внешней среды 1.
Кандиды имеют строение, типичное для всех эукариотических клеток: в цитоплазме присутствуют рибосомы, митохондрии, включения в виде гликогена, крупное ядро, ограниченное ядерной мембраной, аппарат Гольджи, эндоплазматическая сеть.
Клеточная стенка является существенной структурой, обеспечивающей жизнедеятельность кандид, в частности С.аlbicans, и ее взаимодействие с тканями хозяина (рис. 1, а). Она состоит главным образом из углеводов (80-90%), которые представлены бета-глюканом, хитином и маннаном. Компоненты клеточной стенки дрожжевой и гифальной форм сходны по химическому составу, хотя имеют некоторые количественные вариации.
 |
| Рис. 1. Молекулярное взаимодействие между клеточной стенкой C. аlbicans и поверхностями оральной полости (по Cannon, 1999): a - схематическое изображение строения и химического состава клеточной стенки C. аlbicans; б - взаимодействие C. аlbicans с молекулами и поверхностями оральной полости, которые могут подвергаться колонизации
|
Бета-глюкан (разветвленный полимер, содержащий связи b-1,3 и b-1,6) – является главным компонентом клеточной стенки (от 47 до 60% сухой массы). В свою очередь, хитин (неветвящийся полимер N-ацетилглюкозамина) является минорной составляющей клеточной стенки (от 1 до 10% сухой массы). В гифах отмечают более высокий уровень содержания хитина – примерно в 3 раза больше, чем в дрожжевой форме. Эти два микрофибриллярных полисахарида, хотя и распределены по всей клеточной стенке, но более сконцентрированы вблизи плазматической мембраны, обеспечивая ригидность каркаса. Следующим важным компонентом клеточной стенки является маннан (до 40% сухого вещества клеточной стенки), также иногда называемый фосфопептидо-маннановым комплексом. Маннаны состоят из полимеров маннозы, которая может быть ковалентно связана с различными белками через гликозидную связь: ди-N-ацетилхитобиозу и аспарагиновый участок белка 5.
Электронная микроскопия показала вариабельность числа слоев клеточной стенки (от 3 до 8) 21, что зависит от техники посева, штамма и условий выращивания грибов. Эта слоистость может быть результатом различной количественной выраженности главных компонентов клеточной стенки кандид: b-1,3-глюкана, b-1,6-глюкана, хитина, маннопротеина 21. Предполагают, что b-1,6-глюкан с несколькими b-1,3-глюкановыми ветвями может быть прикреплен к восстановленному концу хитина. Ковалентное связывание маннопротеина с b-1,6-глюканом осуществляется через остаток маннопротеина GPI (глюкозил-фосфатидил-инозил). Однако каждый подобный комплекс может содержать не все вышеупомянутые компоненты, а пропорция полисахаридов клеточной стенки, включенных в этот тип структуры, окончательно не известна.
Плазматическая мембрана кандид содержит высокий процент эргостерола и систему энзимов, обеспечивающих его биосинтез. Это химическое своеобразие позволяет использовать мембрану как мишень для антифунгальной терапии. На поверхности клетки находятся фимбрии, длиной от 110 до 300 нм, содержащие высокогликозилированный гликопротеин 22.
Метаболизм кандид
По типу дыхания кандиды являются аэробами и способны метаболизировать глюкозу по гексозо-монофосфатному пути или по пути Эмбдена-Мейергофа-Парнаса (гликолиз). Другие метаболические механизмы, такие как митохондриальное окислительное фосфорилирование, цитратный цикл и синтез белка на рибосомах, не отличаются от таковых в других эукариотических организмах 1,20. Энзиматический аппарат кандид довольно сложен. Многие ферменты, такие как протеазы, обеспечивающие синтез стеролов, могут прямо участвовать в патогенезе заболевания.
Культуральные свойства
Кандиды лучше растут на специальных питательных средах с добавлением углеводов (глюкозы), но могут размножаться и на простых. Оптимальная температура роста грибов – +22-36оС; pH – 5,8-6,5. Специальными средами для выращивания кандид являются глюкозо-пептонный агар, кукурузный питательный агар, среда Сабуро. Колонии на глюкозо-пептонной среде при +25оС через 1-3 дня роста – молочно-белые, беловато-кремовые с тусклым блеском; вначале гладкие, влажные, при дальнейшей инкубации могут стать морщинистыми 1, 2, 4.
Факторы патогенности
Обсуждая патогенетические аспекты кандидоза, надо иметь в виду, что некоторые виды кандид, в частности С.аlbicans, являются комменсалами человека, поэтому кандидозная инфекция носит преимущественно эндогенный характер 3, 19, 23. Колонизация слизистых кандидами может закончиться приобретением и сохранением стабильной популяции кандид, которая не дает развития клинической инфекции. Колонизация зависит от интенсивности приобретения инфектанта, т.е. скорости, с которой клетки кандид проникают в полость рта или другую экологическую нишу, от способности микроорганизмов к адгезии и наличия различных механизмов очищения (клиренса) слизистых. В упрощенной модели это можно представить следующим образом: если скорость удаления больше, чем скорость приобретения и роста кандид, наступает очищение. Если скорость проникновения и удаления одинаковы, может быть колонизация. Если скорость удаления низкая и есть повреждения ткани, то может развиться кандидоз 3. Клинические проявления заболевания могут зависеть от состояния колонизируемой ткани, факторов вирулентности конкретного штамма кандид и ответной защитной реакции хозяина.
Факторы патогенности кандид можно условно разделить на пять групп, хотя в организме, при возникновении патологии, их воздействие осуществляется одновременно 1, 5, 24, 25.
Способность грибов к адгезии на тканях хозяина – первый шаг к взаимодействию с макроорганизмом. В данный процесс вовлечены разнообразные адгезины кандид и рецепторный аппарат слизистых оболочек организма хозяина.
Продукция протеолитических энзимов – секреторных аспартил-протеаз (SAP), облегчающих пенетрацию и инвазию кандид в ткани.
Морфологическая трансформация: дрожжевая-гифальная форма (yeasts-hyphae), которая также способна облегчить кандидам проникновение в ткани и помогает микроорганизму обходить защитные системы хозяина.
Различные иммуномодуляторные действия (механизмы) некоторых поверхностных молекул С.аlbicans, содействующие снижению активности защитных сил хозяина.
Фенотипические переключения, характерные для отдельных штаммов С.аlbicans при изменении условий существования.
Заметим, что в настоящее время наиболее изученными остаются факторы патогенности С.аlbicans.
1. Адгезины и механизмы адгезии. Начальным этапом колонизации является адгезия, которая реализуется через разнообразные механизмы распознавания патогеном (грибом) тканей хозяина. Адгезины – участки поверхности кандид, участвующие в прикреплении последних к клеткам хозяина (эпителиоцитам), микроорганизмам, инертным полимерам и отдельным белкам биологических жидкостей (например, слюны). С.аlbicans обладают множеством адгезинов, имеющих разное химическое строение, кроме этого, для кандид характерно наличие набора адгезинов, участвующих в распознавании отдельного лиганда или клетки хозяина (см.таблицу).
С.аlbicans способна прикрепляться к разным субстратам (рис.1,б), таким как эпителиоциты слизистых (буккальные, вагинальные, дермальные и др.), эндотелиоциты 26, 27, белки слюны, бактерии нормальной микрофлоры и инертные поверхности (различные полимеры, используемые для постоянных медицинских процедур – аппараты, приборы, катетеры) 28-30.
| Механизмы адгезии кандид (по R.D.Cannon et al., 1995) |
| Механизм адгезии |
Адгезины кандид |
Рецептор/лиганд |
Ткани/поверхности хозяина |
| Гидрофобность |
Поверхностные белки/протеины |
Гидрофобные поверхности |
Эпителиальные клетки, зубные материалы |
| Протеин-протеин |
"Интегриноподобные" поверхностные протеины, например, бМв2, бХв2, б5в1 |
iC3b-; C3d-; RGD, содержащие протеиды* (например, фибронектин) |
Эпителиоциты, внеклеточный матрикс, эндотелиальные клетки, кровь |
| Поверхностные протеины |
C3d; внеклеточные матриксные протеины (например, фибриноген, коллаген, ламинин, фибронектин, энтактин) |
Эпителиоциты, внеклеточный матрикс, эндотелиальные клетки, кровь |
| Лектиноподобные |
Поверхностные гликопротеиды |
Фукозные или N-ацетил-D-глюкозаминовые остатки |
Эпителиальные клетки |
| Поверхностные полисахариды |
Полисахариды оральных стрептококков |
Колонизированные эпителиальные клетки |
| Фимбрии-маннопротеины |
Гликосфинголипидные рецепторы |
Эпителиальные клетки |
| Неопределенные |
Не известны |
Гликосфинголипиды |
Эпителиальные клетки |
| Углеводные части маннопротеинов клеточной стенки |
Не известны |
Макрофаги |
| * Полипротеиды, содержащие последовательность аргинин-глицин-аспарагин |
Адгезия в системе «макро-микроорганизм» зависит от условий внешней среды, действующей через кандиды, с одной стороны, и влияющей опосредованно через организм хозяина – с другой. К кандидозависимым механизмам, воздействующим на эту систему, относят: гидрофобность поверхности гриба, тип питательной среды и условия выращивания. В свою очередь, на адгезивные потенции клеток макроорганизма влияет гормональный и иммунный статус хозяина 31-35.
Прикрепление кандид к клеткам хозяина инициирует колонизацию и инфекционный процесс. Эта концепция легла в основу идеи предотвращать развитие инфекции путем блокады адгезии кандид к тканям хозяина и/или ингибированием рецепторного аппарата эпителиоцитов 1, 24. Такой подход может быть новой профилактической моделью для защиты от кандидозной инфекции 24.
Существует мнение, что степень адгезии коррелирует с патогенностью кандид для человека и животных. Среди кандид обнаружены высоко– и низкоадгезивные виды. Наибольшую «активность» в модельных экспериментах на корнеоцитах проявляла С.аlbicans, которая адгезировалась в большем количестве, чем С.tropicalis, в то время как С.krusei показывала незначительную или нулевую адгезию 1.
В процессе адгезии играют роль как неспецифические (гидрофобные контакты), так и специфические механизмы (лиганд-рецепторные взаимодействия). Адгезинами кандид могут быть поверхностные белки, интегриноподобные поверхностные протеины (aMb2, aXb2, a5b1), молекулы, участвующие в лектиноподобных контактах, а также фимбрии. Большинство адгезинов определены как маннопротеины, при этом адгезивными свойствами главным образом обладает их белковый компонент. В некоторых случаях в процессах адгезии участвуют углеводные части маннопротеинов клеточной стенки кандид. Выдвинуто предположение, что фактор-6 маннана (антиген) может быть связан с активностью адгезии 25, 36, 37. Строение соответствующих рецепторов (к адгезинам кандид) на клетках хозяина, по-видимому, зависит от типа клетки и наличия на ее поверхности фукозила, глюкозамина, фибронектина или аргинин-глицин-аспарагина (RGD) 38. Мутантные штаммы, дефицитные по фактору-6 (штаммы серотипа В), проявляют сниженную адгезивность по сравнению с немутантными штаммами (серотип А) 36. Данный факт, возможно, имеет практическое значение: в клинических материалах чаще встречается серотип А, чем серотип В, кроме того, от больных с подавленным иммунитетом чаще выделяется серотип В. При назначении антифунгального лечения необходимо помнить, что серотип А чувствителен к 5-флуороцитозину, а серотип В – часто устойчив 1.
Отметим, что кандиды нередко используют опосредованные механизмы адгезии, так, например, в процесс закрепления грибов на буккальных эпителиоцитах могут вовлекаться молекулы слюны или происходит коадгезия с оральными бактериями-комменсалами. Имеются данные о том, что клетки С.аlbicans могут коадгезироваться с отдельными видами оральных бактерий, включая Streptococcus gordonii, S.mutans, S.oralis, S.sanguis, S.salivarius, Actinomyces species, а пролин-содержащие белки слюны способствуют адсорбции кандид на различных поверхностях ротовой полости 5, 39, 40.
Отдельные виды кандид, такие как С.аlbicans и С.stellatoidea, подобно клеткам человека, способны экспрессировать рецепторы для связывания с iС3b и С3d-молекулами – продуктами расщепления С3-компонента комплемента. Другие виды рода Candida не обнаруживают подобной рецепторной активности. Экспрессия iС3b-рецептора отмечается у различных морфологических форм С.аlbicans, в то время как С3d-рецепторы присутствуют только на гифальных элементах 41-44. iС3b– и С3d-рецепторы С.аlbicans относятся к семейству b2-интегринов (семейство клеточных поверхностных молекул, ответственных за адгезию в системах клетка-клетка и клетка-матрикс) и состоят из трансмембранных гетеродимеров с двумя ковалентно несвязанными a– и b-частями. Установлено, что iС3b-рецептор С.аlbicans обладает структурным сходством с
a-цепью CR3-рецептора клеток человека 18, 42. iС3b-рецептор способен обеспечивать адгезию грибов к эпителиальным и эндотелиальным клеткам 5, 25, что дает возможность предполагать его активное участие в патогенезе кандидозной инфекции.
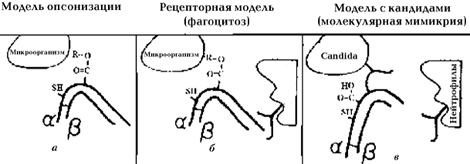 | | Рис. 2. Схематическое изображение механизма, при помощи которого рецепторы кандид ковалентно связывают iC3b и предотвращают распознавание последнего CR3-рецепторами нейтрофилов (по B.J. Gilmore, 1998): а - iC3b ковалентно прикрепляется к микроорганизму (механизм опсонизации); б - в этой форме iC3b способен распознаваться рецепторами нейтрофилов; в - iC3b нековалентно прикрепляется к рецептору кандид, в этом случае участки распознавания для CR3-рецепторов нейтрофилов замаскированы |
Кроме того, iС3b– и C3d-рецепторы кандид обладают функциональной аналогией с соответствующими рецепторами фагоцитарных клеток человека (CR1, CR2, CR3 и CR4) 25, 42, 43. Подобная «мимикрия» обеспечивает конкуренцию между рецепторами кандид и фагоцитами за связывание специфических участков iC3b– и C3d-молекул системы комплемента (рис. 2). Нековалентный «захват» рецепторами кандид iC3b– и C3d-молекул блокирует процесс опсонизации этих микроорганизмов, что снижает эффективность фагоцитоза.
Таким образом, наличие у С.аlbicans и С.stellatoidea iС3b– и C3d-рецепторoв представляется новым атрибутом вирулентности и указывает на то, что эти низшие эукариоты могут подражать белкам млекопитающих для эффективного избегания механизмов защиты хозяина. Изучение адгезивных молекул кандид, в частности, обеспечивающих мимикрию, создает основу для разграничения штаммов по вирулентности и выявления потенциальных патогенов 41.
2. Продукция протеолитических энзимов. После начального этапа взаимодействия между дрожжеподобными клетками и эпителиоцитами хозяина начинают подключаться факторы, участвующие в проникновении (пенетрации) и распространении (инвазии) кандид в ткани. Пенетрация и инвазия обеспечиваются морфологическими изменениями от дрожжеподобных форм к гифам (см. ниже) и продукцией гидролитических ферментов 1. Клетки C.albicans секретируют фосфолипазу, липазу, фосфомоноэстеразу, гексозаминидазу и протеолитические энзимы – аспартил-протеазы (SAP) 3, 45, 46. Показано, что экспрессия экзоферментов-протеиназ зависит от штамма, морфологии клеток и факторов окружающей среды 47.
Имеются данные, что аспартил-протеазы содействуют процессу адгезии. В частности, они способны воздействовать на поверхность дрожжей, модифицируя их адгезины, или на клетки хозяина, обнажая лиганды 47. Доказано, что их продукция может облегчить инвазию посредством разрушения кератина и коллагена 1.
SAP-продукция доказана у патогенных видов Сandida: выраженная энзимная активность обнаружена у C.albicans, C.tropicalis, C.parapsilosis. Другие виды кандид лишены этой активности или проявляют ее только частично. При экспериментальной инфекции SAP-дефицитные мутанты были менее патогенны и не способны к инвазии в ткани 1. Молекулярные и генетические исследования аспартил-протеаз у С.аlbicans показали, что существуют несколько видов, каждый из которых кодируется отдельным геном 45. Кроме того, С.аlbicans может продуцировать гемолитические факторы 48, 49. Полагают, что подобное свойство кандид способно в ряде случаев облегчить доступ грибам к железу через поврежденную поверхность эритроцитов.
3. Морфологическая трансформация: дрожжевая – гифальная форма (yeasts – hyphae трансформация). Обнаружение инвазии кандид в ткани служит критерием инфекционного процесса 1. В то же время для носительства характерно присутствие грибов на поверхности эпителия. Инвазия кандид в тканевые структуры осуществляется благодаря механическим и ферментным факторам и сопровождается морфологической трансформацией из дрожжевой в гифальную форму 50. Морфогенез дрожжей в гифальные элементы может быть отнесен к факторам патогенности 1, 3, поскольку наблюдается только при активной кандидозной инфекции 39. На трансформацию кандид оказывает влияние температурный режим. Так, диапазон от 37 до 40оС (температура организма потенциальных больных) содействует образованию гифальных форм.
Феномен морфологической трансформации, тесно связанный с патогенностью, приводит к реорганизации клеточной стенки на молекулярном уровне и, как результат, к увеличению спектра адгезинов у гифальных элементов. Важны, по крайней мере, два изменения в клеточной стенке для приобретения гифами более выраженных адгезивных свойств: развитие фибриллярного маннопротеинового покрытия, связанного с внешней поверхностью клеточной стенки, и наличие специфических маннопротеинов. Кроме того, мицелиальная трансформация, как указывалось выше, коррелирует с экспрессией рецепторов к C3d-молекулам и усилением экспрессии iC3b-рецепторов, что значительно увеличивает резистентность гифальных форм к фагоцитозу 41.
Формирование гиф облегчает миграцию кандид через поврежденные ткани. Гифы способны к тигмотропизму – движению, стимулированному чувствительным контактом 51. Гифы в тканях распространяются быстрее, чем дрожжевая форма, что также способствует миграции кандид через поврежденные ткани и пенетрации в здоровые ткани 5. Механизм инвазии может быть представлен следующим образом: вначале адгезия осуществляется между дрожжеподобными клетками кандид и поверхностью эпителиоцитов. Затем под влиянием гидролитических ферментов (в частности, аспартил-протеаз) формируются полости в месте прикрепления клетки гриба; начинается трансформация и продуцируются гифы, которые поражают эпителиоциты дистальнее прикрепления дрожжевых форм 52. Фосфолипазы, концентрируясь на концах гиф, обусловливают большую инвазивность этой формы по сравнению с дрожжевой 10, 53. Гифальные элементы – крупнее, чем дрожжевая форма, и лишены маннановых адгезинов для контакта с макрофагами 3, 54, 55, что служит дополнительными атрибутами устойчивости гиф к фагоцитозу. Таким образом, гифальная форма является более агрессивной и патогенной 1. Однако нет окончательных доказательств того, что только она является вирулентной для макроорганизма, поскольку гистологические исследования кандидозных повреждений иногда показывают отсутствие гиф в патологическом материале 3, 52.
4. Иммуномодуляторные механизмы некоторых поверхностных молекул кандид, содействующие снижению активности защитных сил хозяина. Некоторые поверхностные компоненты кандид, по-видимому, обладают иммуномодуляторной активностью. Так, супрессивное действие на Т-зависимые иммунные реакции главным образом связывают с маннаном клеточной стенки 56-58. В свою очередь, как упоминалось выше, рецепторы кандид к iC3b– и C3d-факторам комплемента способны модулировать фагоцитарный ответ 25, 42, 44, 55.
5. Фенотипические переключения. Дополнительным атрибутом вирулентности является фенотипическое переключение, отмечаемое у отдельных штаммов С.аlbicans 59. Это переключение делает возможной адаптацию к различным и/или изменяющимся условиям и, таким образом, помогает кандидам преодолевать защитные механизмы хозяина. Были обнаружены гены ALS-семейства, которые отвечают за процессы адгезии, и установлено, что контакт C.albicans с эпителиоцитами способен вызывать изменение экспрессии подобных генов 60. При этом может быть изменена степень адгезии, которая коррелирует с патогенностью Candida spp. Варианты внутри видов также могут обладать разной способностью к адгезии 61. Известно, что мутантные штаммы кандид со сниженной адгезивностью проявляют пониженную патогенность in vivo 62. Кроме того, имеются данные о том, что у авирулентных мутантов C.albicans редуцируется способность к iC3b– рецепции 63.
Феномен фенотипического переключения рассматривается как аналогия «фазовой трансформации» у бактерий. Установлено, что «master switch» (переключатель) ответственен за выключение одной группы генов и включение других групп, некоторые из которых определяют вирулентность. Контакт со специфической поверхностью активирует группу генов, обеспечивающих синтез факторов, задействованных в адгезии и пенетрации 5. У некоторых штаммов С.аlbicans обнаружена морфологическая трансформация колоний под влиянием питания, что сопровождается изменением морфологии клеток и в ряде случаев – хромосомной транслокацией 64, 65. Высокочастотное фенотипическое переключение способно одновременно воздействовать на проявление многих потенциальных факторов вирулентности и может быть генетическим механизмом, позволяющим различным формам С.аlbicans адаптироваться к изменению среды.
Заключение
Таким образом, многие виды кандид, в частности С. аlbicans, являются частью нормальной микрофлоры слизистых оболочек. При развитии заболевания в патогенез кандидозной инфекции вовлекаются как факторы патогенности кандид, так и система защиты организма хозяина. Главным вопросом остается определение «пусковых» механизмов, приводящих к нарушению баланса в системе кандиды-слизистые оболочки и преодолению грибами механизмов клиренса.
Кандиды используют разнообразные приемы, позволяющие им адгезировать и колонизировать ткани. Процесс от адгезивной репликации грибов до перехода к инфекциям слизистых оболочек зависит не только от эффективности адгезии и скорости роста дрожжевых форм, но также и от факторов, обеспечивающих пенетрацию в ткани. Выделение гидролитических ферментов, трансформация дрожжевых клеток в гифальные элементы, способность к тигмотропизму – все эти факторы облегчают данный процесс. В свою очередь, поводом для развития инфекции служит и то, что иммунная система человека не в состоянии сдерживать быструю и массивную атаку кандид. Баланс между механизмами очищения, колонизацией и развитием кандидозной инфекции зависит от способности грибов рода Candida модулировать экспрессию факторов вирулентности в ответ на изменение окружающей среды в сочетании с компетентностью иммунной системы хозяина.
Литература
Topley and Wilson`s. Microbiology and Microbial Infections. Vol. Medical Mycology. 9th ed.1999 (Online version).
Антоньев А.А., Бульвахтер Л.А., Глазкова Л.К., Ильин И.И. Кандидоз кожи и слизистых оболочек. М: Медицина; 1985; 155 с.
Cannon R.D., Holmes A.R., Mason A.B., Monk B.C. Oral candida: clearance, colonization, or candidiasis?J Dent Res 1995; 74(5): 1152-1161.
Бурова С.А., Воинова Г.В. Клинические разновидности и лечение кандидоза. Вестн дерматол 1997; 4: 24-28.
Cannon R.D., Chaffin W.L. Oral colonization by Candida albicans. Crit Rev Oral Biol 1999; 10 (3): 359-383.
Мюллер Э., Лёффлер В. Микология. М: Мир; 1995; 343 с.
Magee P.T. and Stewart Scherer. Genome Mapping and Gene Discovery in Candida albicans. ASM News 1998; 64 (9): 505-511.
Реброва Р.Н. Грибы рода Candida при заболеваниях негрибковой этиологии. М: Медицина; 1989;128 с.
Rangel-Frausto M. S., Houston A.K., Bale M.J., Fu C., Wenzel R.P. An experimental model for study of Candida survival and transmission in human volunteers. Eur J Clin Microbiol Infect Dis 1994; 13: 590-595.
Odds F.S. Candida and candidosis. 2nd ed. London: Bailliere Tindall; 1988.
Cole G.T., Halawa A.A., Anaissie E.J. The role of gastrointestinal tract in hematogenous candidiasis: from the laboratory to the bedside. Clin Infect Dis 1996; 22: 73-88.
Pawderly W.G., Robinson R., Keath E.J. Molecular epidemiology of recurrent oral candidiasis in human immunodefency virus-positive patients: evidence for two patterns of recurrence. J Infect Dis 1993; 168: 463-466.
Voss A., Hollis R.J., Pfaller M.A., Wenzel R.P., Doebble B.N. Investigation of the sequence of colonization and candidemia in nonneutropenic patients. J Clin Microbiol 1994; 32: 975-980.
Schmid J., Voss E., Soll D.R. Computer-assisted methods for assessing strain relatedness in Candida albicans by fingerprinting with the moderately repetitive sequence Ca3. J Clin Microbiol 1990; 28(6):1236-1243.
Vazquez J.A., Sancher V., Dmuchowski C., Dembry L.M., Sobel J.D., Zervos M. J. Nosocomial acquisition of Candida albicans: an epidemiologic study. J Infect Dis 1993;168 (1):195-201.
16. Arendorf T.M., Walker D.M. Oral candidal polulations in health and disease. Br Dent J 1979; 147: 267-272.
Viljoen B.C., Greyling T. Yeast associated with Chedda and Gouda making. Int S Food Microbiol 1995; 28: 79-88.
Pendrak M.Z., Klotz S.A. Adherence of Candida albicans to host cells. FEMS Microbiology letters 1995; 129: 103-114.
Самсыгина Г.А., Буслаева Г.Н., Корнюшин М.А. Кандидоз новорожденных и детей раннего возраста. Дифлюкан в лечении и профилактике кандидоза. М; 1996; 40 с.
Braude A. Candida, Infectious diseases and Medical Microbiology. 2nd ed. 1986; 571р.
Rico H., Herrero E., Miragall F., Sentandreu R. An electron microscopy study of wall expansion during Candida albicans yeast and mycelial growth using concanavalin A-ferritin labelling of mannoproteins. Arch Microbiol 1991; 156: 111-114.
Yu Z., Zee K.K., Ens K. Doig P.C., Carpenter M.R., Staddon W. Partial characterization of a Candida albicans fimbrial adhesin. Infect Immun 1994; 62: 2834-2842.
Прилепская В.Н., Анкирская А.С., Байрамова Г.Р., Муравьева В.В. Вагинальный кандидоз. М; 1997;
40 с.
Segal E. Inhibitors of Candida albicans adhesion to prevent candidiasis. Adv Exp Med Biol 1996; 408: 197-206.
Calderone R.A., Braun P. Adherence and receptor relationship of Candida albicans. Microbiol Rev 1991; 55:1-20.
Gustafson K.S., Vercellotti G.M., Bendel C.M., Hostetter M.K. Molecular mimicry in Candida albicans. J Clin Invest 1991; 87: 1896-1902.
Segal E., Gottlieb S., Altboum Z., Goy V., Berdicevsky J. Adhesion of Candida albicans to epithlial cells- effect of nikkomycin. Mycoses 1997; 40: 33-39.
Chaffin W.Z., Lopez-Ribot J.Z., Casanova M., Gozalbo D., Martinez J.P. Cell wall and secreted proteins of Candida albicans: identification, function and expression. Microbiol Mol Biol Rev 1998; 62: 130-180.
Van Weissenbruch R., Bouckaert S., Remon J.P., Nelis H.J., Albers F.W. Chemoprophylaxis of fungal deterioration of the Provox in post-laryngectomy patients. Ann Oto Rhinol Laryngol. 1997; 106: 329-337.
Busscher Y.S., Geertsema-Doornbusch G.G., van der Mei H.C. Adhesion to silicone rubber of yeast and bacteria isolated from voice prostheses: influence of salivary conditioning films. J Biomed Mat Res 1997; 34: 201-209.
Sturtevant J., Calderone R. Candida albicans adhesins: biochemical aspects and virulence. Rev Jberoam Micol 1997; 14: 90-97.
Theaker E.D., Drucker D.B. et al. The possible influence of the menstrual cycle of the adherence of Candida albicans to human buccal epithelial cell in vitro. Arch Oral Biol 1993; 38: 353-355.
Segal E., Soroka A., Schecter A. Correlative relationship between adherence of Candida albicans to human vaginal epithelial cells in vitro and candidal vaginitis. Sabouraudia 1984; 22: 191-200.
Wellmer A.,Berhardt H. Adherence on buccal epithelial cells and germ tube formation in the continuous flow culture of clinical Candida albicans isolated . Mycoses 1997; 40: 363-368.
Pereiro M., Losada A., Toribo S. Adherence of Candida albicans strains isolated from AIDS patients. Comparison with pathogenic yeasts isolated from patients without HIV infection. Br J Dermatol 1997; 137: 76-80.
Bendel C.M., Hostetter M.K. Distinct mechanisms of epithelial adhesion for Candida albicans and Candida tropicalis. JClin Invest 1993; 92: 1840-1849.
Miyakawa V., Kuribayashi T., Kagaya K. Role of specific determinant in mannan of Candida albicans serotype A in adherence to human buccal epithelial cells. Infect Immun 1992; 60: 2493-2499.
Hostetter M.K. Adhesins and ligands involved in the interaction of Candida spp. With epithelial and endothelial surfaces. Clin Microbiol Rev 1994; 7: 29-42.
Nair R.G., Samaranayake L.P. The effect of oral commensal bacteria on candidal adhesion to human buccal epithelial cells in vitro. J Med Microbiol 1996; 45: 179-185.
O`Sullivan J.M., Jenkison H.F., Cannon R.O. Adhesion of Candida albicans to oral streptococci in promoted by selective adsorption of salivary proteins to the streptococcal cell surface. Microbiology 2000; 146: 41-48.
Tronchin G., Bouchara J.P., Annaix V., Robert R. D., Senet J. M. Fungal cell adhesion molecules in Candida albicans. Eur J Cell Biol 1991; 7(1): 23-33.
Gilmore B.J., Retsinas E.M., Lorenz J.S., Hostetter M.K. An iC3b receptor on Candida albicans: structure, function and correlates for pathogenicity. J Infec Dis 1988. 157 (1): 38-46.
Кашкин К.П., Дмитриева Л.Н. Белки системы комплемента: свойства и биологическая активность. Клин лаб диагн 2000; 7: 25-32.
Challacombe S.J. Immunologic aspects of oral candidiasis. Oral Surg Oral Med Oral Pathol 1994; 78 (2): 202-210.
White T.C. Agabian N. Candida albicans secreted aspartyl proteinases: isoenzyme pattern is determined by cell type, and levels are determined by environmental factors. J Bacteriol 1995; 177: 5215-5221.
Odds F.C. Candida species and virulence. ASM News 1994; 60: 313-318.
Watts H.J., Cheah F.S.H., Yube D., Sanglard D., Gow N.A.R. Altered adherence in strains of Candida albicans harbouring null mutations in secreted aspartic proteinase genes. FEMS Microbiol Lett 1998; 159: 129-135.
Manns J.M., Mosser D.M., Buckley H.R. Production of a hemolytic factors by Candida albicans. Infect Immun 1994; 62: 5154-5156.
Караев З.О., Лебедева Т.Н. К вопросу о биологической активности Candida albicans. В кн.: Микотическая инфекция и сенсибилизация. Сборник науч. труд. Ленинград; 1982; с. 14-17.
-
Быков В.Л. Динамика инвазивного роста Candida albicans в тканях хозяина. Вест дерматол венерол 1990; 4: 25-28.
Sherwood J., Gow N.A.R., Gooday G.M., Gregory D.M., Marshall D. Contact sensing in Candida albicans: a possible aid to epithelial penetration. J Med Vet Mycol 1992; 30: 461-469.
Edwards J.E. J., Mayer C.L., Filler S.D., Wadsworth E., Calderone R.A. Cell extracts of Candida albicans block adherence of the organisms to endothelial cel. Infect Immun 1992; 60: 3087-3091.
Ponton J., Bikandi J., Moragues M.D., Arilla M.C., Elosegui R., Quindos G. Reactivity of Candida
albicans germ tubes with salivary secretory IgA. J Dent Res 1996; 75: 1979-1985.
Diamond R.D. Interaction of phagocytic cells with Candida and other opportunistic fungi. Arch Med Res 1993; 24: 361-369.
Cassone A. Cell wall of Candida albicans: its functions and its impact on the host. Curr Top Med Mycol 1989; 3: 248-314.
Domer J.E., Stashak P.W., Elkins K. Prescott B., Caldes G., Baker P.J. Separation of immunomodulatory effects of mannan from Candida albicans into stimulatory and suppressive components. Cell Immunol 1986; 101 (2): 403-414
Garner R.E., Childress A.M., Human L.G., Domer J.E. Characterization of Candida albicans mannan-induced, mannan-specific delayed hypersensitivity suppressor cells. Infect Immun 1990, 58 (8): 2613- 2620.
Soll D.R. High-frequency switching in Candida albicans. Clin Microbiol Rev 1992; 5 (2): 183-203.
Bailey A., Wadsworth E., Calderone R. Adherence of Candida albicans to human buccal epithelial cells: host-induced protein synthesis and signaling events. Infect Immun 1995; 63 (2): 569-572.
Samaranayake Y.H., Wu P.C., Samaranayake L.P., So M. Relationship between the cell surface hydrophobicity and adherence of Candida krusei and Candida albicans to epithelial and denture acrylic surfaces. APMIS 1995; 103 (10): 707-713
Calderone R.A., Cihlar R.L., Lee D.D., Hoberg K., Scheld W.M. Yeast adhesion in the pathogenesis of endocarditis due to Candida albicans: studies with adherence-negative mutants. J Infect Dis 1985; 152 (4): 710-715.
Ollert M.W., Wadsworth E., Calderone R. A. Reduction expression of the functionally active complement receptors for iC3b but not C3d on an avirilent mutant of Candida albicans. Infect Immun 1990; 58 (4): 909-913.
Bergen M.S., Voss E., Soll D.R. Switching at the cellular level in the white- opaque transition of Candida albicans. J Gen Microbiol 1990; 136 (10): 1925-1936.
Srikantha T., Morrow B., Schroppel K., Soll D.R. The frequency of integrative transformation at phase-specific genes of Candida albicans correlates with their transcriptional state. Mol Gen Genet 1995; 246 (3): 342-35265.
|
Поможем с водопотреблением и водоотведением https://ecopromcentr.ru/ для предприятий.
Вверх | Назад
Главная |
Врачам |
Пациентам |
Студентам |
Мед.учреждения |
Мед.анекдоты |
Полезные ссылки
Нижегородский медицинский сайт
по вопросам размещения рекламы пишите здесь
|
|