© Н.Н. Андреева, И.В. Мухина, Т.И. Соловьева,
Е.К. Слюняева, 2003 г.
УДК 612.111.15:616.831
Поступила 30.06.2003 г.
Н.Н. Андреева, И.В. Мухина, Т.И. Соловьева, Е.К. Слюняева
Центральная научно-исследовательская лаборатория НГМА, Нижний Новгород
Изменения спектрального состава липидов мозга в постреанимационном периоде
Известно, что реоксигенация и рециркуляция в организмах, переживших терминальное состояние (тяжелую гипоксию в результате остановки кровообращения), не только обеспечивают восстановление функций ЦНС, но и вызывают новые нарушения метаболизма мозга, оказывающие значительное влияние на течение и исход постишемического процесса [1, 2]. Одно из наиболее существенных биохимических нарушений, характерных именно для реперфузионного периода, – это нарушение обмена фосфолипидов, основного компонента мембран клеток [3]. Гомеостаз мембран определяется тремя факторами: синтезом фосфолипидов de novo; фосфолипазами и ацил-трансферазами, осуществляющими реакции деацилирования/реацилирования; процессами перекисного окисления липидов (ПОЛ). Фосфолипиды, образуя структурную основу биомембран и определяя ее микровязкость, выполняют роль эффекторов липидзависимых ферментов, обеспечивающих метаболические, транспортные, регуляторные и рецепторные функции [4]. Поэтому преобладание процессов деструкции фосфолипидного компонента мембран, нарушение их гомеостаза при ишемии является одним из препятствий восстановления активности липидзависимых мембранных ферментов Na+, К+, АТФазы, Са2+-АТФазы, аденилатциклазы, ферментов дыхательной цепи и, следовательно, полноценного восстановления функций клеток в реперфузионном периоде [3, 4]. Несмотря на значимость изменений метаболизма липидов в развитии ишемических и постишемических нарушений мозга, сведения о спектральном составе фосфолипидов мозга в отдаленном постреперфузионном периоде практически отсутствуют. В связи с этим представляет большой интерес исследование спектра фосфолипидов мозга в различные сроки постишемического периода. Материалы и методы. Эксперименты выполнены на белых нелинейных крысах-самцах массой 180—220 г, содержащихся на стандартном рационе вивария. Для наркоза использовали нембутал (25 мг/кг, внутрибрюшинно) и эфир. Тотальную ишемию моделировали путем пережатия сердечно-сосудистого пучка [5]. Длительность прекращения кровообращения составила 10 мин. Оживление проводили, применяя непрямой массаж сердца и искусственную вентиляцию легких. Забор мозга у наркотизированных животных осуществляли на 60-й минуте восстановительного периода (n=12), на 14-е сутки (n=8) и на 30-е сутки (n=8) постишемического периода. Контролем служили интактные животные (n=12).
Липиды экстрагировали по J.Folch [6]. Разделение липидов на фракции проводили методом тонкослойной хроматографии [7]. Количественную оценку фракций фосфолипидов и нейтральных липидов осуществляли на сканере Scan Maker E6 ф. «Microtek» с использованием программы «Sigma Gel». Содержание отдельных классов липидов выражали в процентах от суммы площадей пиков, принятой за 100%, как описано в работе[8]. Статистическую достоверность различий оценивали методом Стьюдента. Различие считали достоверным при р Результаты и обсуждение. В ранний реперфузионный период после тотальной ишемии в ткани мозга уменьшилось содержание фосфатидилхолина (ФХ), фосфатидилэтаноламина (ФЭ) и фосфатидилсерина (ФС) на 32, 20 и 47% соответственно. параллельно отмечалось накопление лизоформ ФЭ и ФС, фосфатидных кислот (ФК), диацилглицеридов (ДГ) и увеличение количества свободных жирных кислот (СЖК) на 63% (см. таблицу). Эти значительные изменения спектрального состава липидов свидетельствуют как о нарушении ацилобменного механизма регулирования физических и функциональных свойств мембран клеток, так и об угнетении синтеза фосфолипидов de novo [9, 10]. Уменьшение содержания ФЭ и ФС в фосфолипидном спектре ткани мозга ведет к снижению текучести мембран, которое отчасти компенсируется уменьшением количества холестерина (ХС) – на 32%, оказывающего конденсирующее действие на фосфолипиды [11]. СЖК, лизоФС и лизоФЭ, содержание которых увеличилось вследствие активации фосфолипаз и нарушения динамического равновесия между процессами деацилирования и реацилирования, оказывают повреждающее воздействие на мембраны клеток [3, 9].
В ранний реперфузионный период в ткани мозга усиливается не только фосфолиполиз, но и гидролиз нейтральных липидов, о чем свидетельствует уменьшение количества триацилглицеридов (ТГ) на 39% и эфиров холестерина (ЭХС) на 59%.
Снижение количества лизоФХ на 43% к 60-й минуте постишемического периода в ткани мозга при уменьшении содержания ФХ обусловлено либо участием в фосфолиполизе ФХ обеих фосфолипаз –А1 и А2, либо дальнейшим гидролизом лизоФХ [4, 11].
На 14-е сутки постишемического периода содержание основных мембранных фосфолипидов – ФХ и ФЭ — не нормализовалось: их количество было ниже исходного значения на 37 и 23% соответственно (см. таблицу). Одновременно увеличилось содержание СЖК на 46%, накапливался лизоФЭ. Эти изменения липидного спектра свидетельствуют о доминировании реакций деацилирования фосфолипидов в исследуемый период. Определенный вклад в увеличение СЖК в ткани мозга на 14-е сутки постреперфузионного периода вносит и гидролиз нейтральных липидов, так как содержание ТГ и ЭХС уменьшилось на 34 и 58% соответственно и накапливались ДГ. Известно, что образование фосфолипидов de novo – энергозависимый процесс, и в ряде работ [12, 13] показано, что в реперфузионный период в ткани мозга не происходит полноценного восстановления энергетического статуса клеток. В связи с этим можно предположить, что повышение уровня ДГ и ФК — промежуточных продуктов синтеза фосфолипидов [11] — является следствием неполного восстановления синтеза фосфолипидов. На фоне снижения содержания ФЭ и ФХ уменьшение количества холестерина на 16% и увеличение ФС в 2 раза в исследуемый период по сравнению с интактной серией является защитно-приспособительной реакцией клеток, обеспечивающей регуляцию микровязкости липидного компонента мембран [4, 11]. С другой стороны, увеличение ФС частично нивелирует потерю ФХ и ФЭ.
| Содержание липидов в мозге в постишемическом периоде (М±m), % |
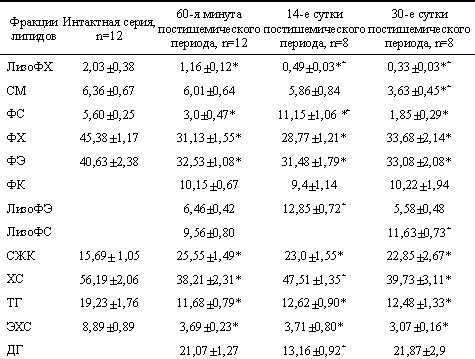 |
* достоверность различий с интактной серией;
+ достоверность различий с ранним постишемическим периодом (60-я минута), р < 0,05. Здесь СМ — сфингомиелин. |
Повышение количества ФС возможно за счет реакций обмена азотистыми основаниями между ФХ, ФЭ и ФС, осуществляемыми ферментами холинспецифической и этанолспецифической фосфатидилсеринсинтетазами [10], и реацилирования лизоФС [4], так как данная фракция в ткани мозга на 14-е сутки по сравнению с ранним постишемическим периодом отсутствовала. Не исключено, что отсутствие лизоФС в это время является результатом действия лизофосфолипаз, так как наблюдалось и уменьшение содержания лизоФХ без изменения количества ФХ. Эти данные свидетельствуют о наличии в этот период и защитно-приспособительных реакций. Однако на 14-е сутки постишемического периода относительно раннего реперфузионного периода не наблюдалось изменения содержания ФХ, ФЭ и СЖК, следовательно, дисбаланс процессов деацилирования/реацилирования фосфолипидов сохраняется.
Значительное увеличение лизоФЭ (в 2 раза) без изменения уровня ФЭ и ФК, а также уменьшение содержания ДГ на 40% при сохранении пула ТГ на 14-е сутки по сравнению с 60-й минутой постишемического периода, свидетельствуют в пользу образования лизоФЭ de novo как промежуточного продукта синтеза ФЭ [4]. Однако образование лизоФЭ без дальнейшей его утилизации в реакциях синтеза ФЭ из-за энергодефицита может оказывать детергентное действие на мембраны клеток [3, 4].
Таким образом, несмотря на развивающиеся адаптационные изменения липидного спектра, на 14-е сутки постишемического периода в мозге превалирует деградация фосфолипидов и не происходит полноценного восстановления механизмов, обеспечивающих гомеостаз мембран.
На 30-е сутки постишемического периода направленность изменений фосфолипидного спектра ткани мозга по сравнению с интактной серией была аналогична изменениям в раннем восстановительном периоде и на 14-е сутки постреперфузионного периода, но они были более выражены.
Так, на 30-е сутки после тотальной ишемии в ткани мозга уменьшилось содержание не только ФХ и ФЭ, но и ФС и СМ на 67 и 43% соответственно, что свидетельствует о более глубокой делипидизации мембран и срыве защитно-приспособительных реакций, имеющих место на 14-е сутки (см. таблицу). Доказательством доминирования в ткани мозга фосфолиполиза в исследуемый период является повышение содержания СЖК на 45% и накопление лизоформ ФЭ и ФС по сравнению с интактной серией. Увеличение количества СЖК обусловлено и активацией липолиза, так как содержание ЭХ и ТГ уменьшилось на 66 и 35% соответственно. На 30-е сутки наблюдалось не только угнетение процессов реацилирования фосфолипидов, но и синтеза их de novo: содержание индивидуальных фосфолипидов не нормализовалось и накапливались ФК и ДГ. Заключение. Таким образом, в постишемическом периоде нарушается гомеостаз мембран клеток мозга, о чем свидетельствуют сохраняющийся длительное время дисбаланс между реакциями деацилирования/реацилирования фосфолипидов, угнетение синтеза фосфолипидов de novo. В связи с вышесказанным считаем целесообразным применение в реперфузионном периоде препаратов мембраностабилизирующего действия для предупреждения развития постгипоксических энцефалопатий.
Литература
- Мутускина Е.А., Заржецкий Ю.В., Трубина И.Е. и др. О возможности уменьшения постреанимационной патологии посредством препаратов янтарной кислоты. В кн.: Экспериментальные, клинические и организационные проблемы общей реаниматологии. Под ред. В. А. Неговского. М: 1996; с. 195—205.
- Неговский В.А., Гурвич А.М. Постреанимационная болезнь – новая нозологическая единица. Реальность и значение. В кн.: Экспериментальные, клинические и организационные проблемы общей реаниматологии. Под ред. В. А. Неговского. М: 1996; с. 3—10.
- Оковитый С.В., Смирнов А.В. Антигипоксанты. Экспер и клин фармакол 2001; 64 (3): 76—80.
- Биленко М.В. Ишемические и реперфузионные повреждения органов. М: Медицина; 1989; 368 с.
- Корпачев В.Г., Лысенков С.П., Тель Л.З. Моделирование клинической смерти и постреанимационной болезни у крыс. Пат физиол и эксперим терапия 1982; 3: 78—80.
- Folch J., Lees M., Stanley G. A simple method for the isolation and purification of total lipids from animal tissues. Biol Chem 1957; 226 (2): 497—509.
- Шаршунова М., Шварц В., Михалец Ч. Тонкослойная хроматография в фармации и клинической биохимии. Т. 2. М: Мир; 1980; с. 536—541.
- Пецев Н., Коцев Н. Справочник по газовой хроматографии. М: Мир; 1987; с. 111—541.
- Туманова С.Ю. Липиды центральной нервной системы и структура клеточных мембран. В кн.: Нейрохимия. Под ред. И. П. Ашмарина, П. В. Стукалова. М: Изд-во Ин-та биомедицинской химии РАМН; 1996; с. 96—144.
- Vance D.E. Glycerolipids biosynthesis in eukaryotes. In: Biochemistry of Lipids, Lipoprotein and Membranes. Amsterdam; 1996; р. 153—180.
- Климов А.Н., Никульчева Н.Г. Обмен липидов и липопротеидов и его нарушения. СПб: Питер; 1999; 505 с.
- Мухина И.В., Бояринов Г.А., Пенкнович А.А. Механизмы действия актовегина на центральную нервную систему в постишемическом периоде. Бюл эксперим биол и мед 1998; 126 (10): 395—398.
- Newnam G.C., Hospod F.E., Trowbridge S.D. et al. Restoring adenine nucleotides in brain slice model of cerebral reperfusion. Cereb Blood Flow Metab 1998; 18 (6): 675—685.